После затянувшейся паузы я хотел было продолжить рассказ о различиях между ноотропами и пcиxocтимyлятopами, но понял, что написать понятное, краткое и содержательное сообщение без разговора о памяти – мишени их действия, которая меня интересует, не получится.
О некоторых молекулярных аспектах памяти я и хочу рассказать.
Но прежде – ответы на вопросы и комментарии к предыдущему посту.
1)На сегодняшний день доказательства усиливающих мышление и память свойств ноотропных препаратов не такие убедительные, как хотелось бы. Я не буду давать ссылки – у меня несколько гигабайтов научной литературы по теме ноотропных препаратов (правда, преимущественно посвящённой одному их классу), так что это физически невозможно. Но я и не ставлю себе задачу (пока) приводить эти доказательства. Хочу, правда, отметить, что в Западной Европе ноотропные препараты (равно, кстати, как и пcиxocтимyлятopы) зарегистрированы в качестве лекарственных средств, хотя, опять-таки, это не означает их эффективность.
2)Некоторые не поняли, в чём состоял смысл прошлого поста :-D. Поясняю. Прежде чем анализировать то или иное явление, ему надо дать определение – вычленить присущие ему черты. Феномену (т.е. явлению) ноотропных препаратов посвящено большое количество информации, полученной серьёзными исследователями, я попытался на её основе привести такое определение, в котором этот класс физиологически активных веществ был бы противопоставлен другому классу веществ – пcиxocтимyлятopам.
3)Многие из вас делились впечатлениями о том, какие эффекты на вас оказывали/не оказывали ноотропные препараты. К сожалению, ваши отдельные случаи не могут быть доказательствами как наличия, так и отсутствия какого-либо эффекта в связи с тем, что а) существует эффект плацебо и б) уровень вариабельности ответов ЦНС на фармакологические вмешательства ощутимо высок, т.е., грубо говоря, эффекты, наблюдаемые у вас, могут совсем не проявиться у вашего коллеги или соседа по лестничной клетке.
А теперь, наконец, поговорим о памяти.
Вынужден вас разочаровать: молекулярный субстрат памяти до сих пор точно не известен. Более того, нет уверенности в расположении этого субстрата: является ли он частью синапса, т.е. места химической передачи нервного импульса между нейронами, или лежит внесинаптически, например в соме (теле) нервной клетке.
Тем не менее, есть немало сведений о каскаде биохимических реакций, которые протекают в ходе обучения и возникновения долговременной памяти.
Несмотря на то, что первая информация о молекулярных механизмах памяти была получена из опытов на беспозвоночных животных, характер процессов в наших с вами мозгах принципиально не отличается. Хотя бы потому, что в них принимают участие те же игроки.
Прежде чем перейти на молекулярный уровень, пару физиологических ремарок.
Вместо того, чтобы давать определение памяти, попрошу вас вернуться на урок литературы, на котором вы изучали «Евгения Онегина», ну или, скажем, «Новую зямлю» Якуба Коласа.
И вот я называю вам строчку «Мой дядя самых честных правил» или «Якi ты, хлопча, нецярпячка», как вы сразу же продолжаете!
На самом деле происходит вот что. Связь между нейронами, хранящими образы Пушкина, Коласа, кабинетов литературы, книг и собственно содержимое произведений настолько сильна, что импульс, предъявляющий первую строчку, сразу и быстро пробегает по цепочке нейронов и вызывает из памяти последующую. Таким образом, память определяется силой связи между нервными клетками. Чем она сильнее, тем прочнее память, так как её содержимое воспроизводится точнее и быстрее.
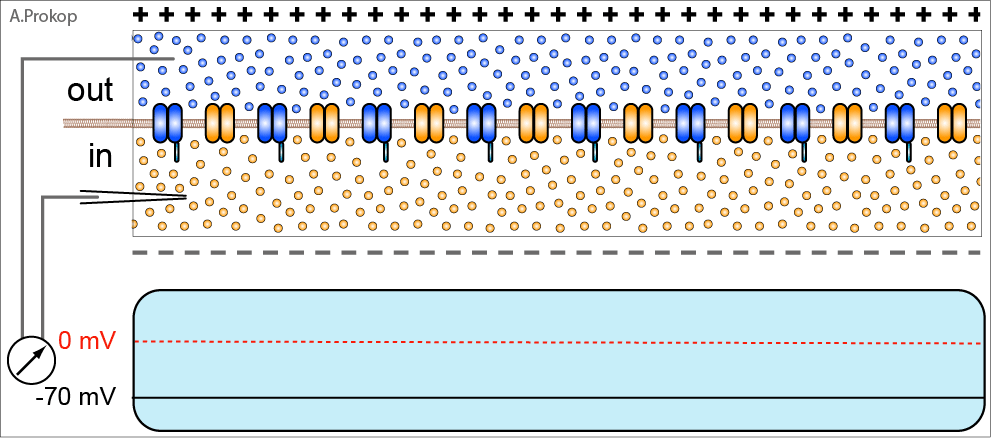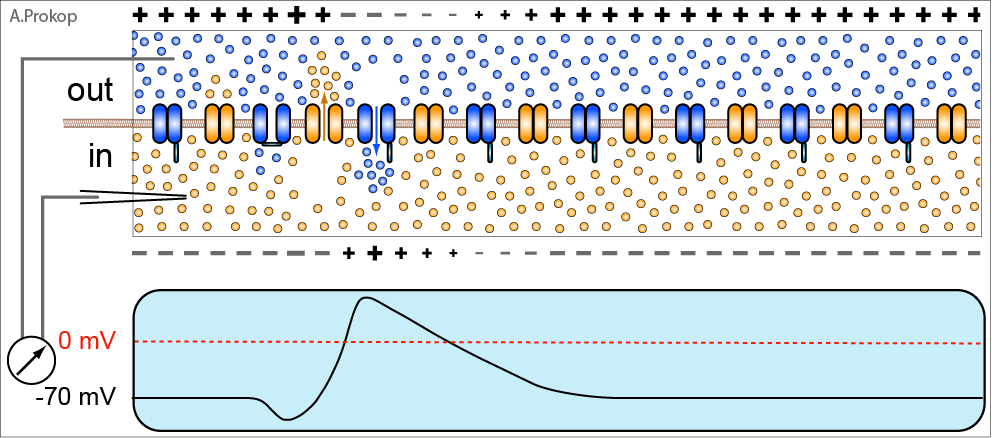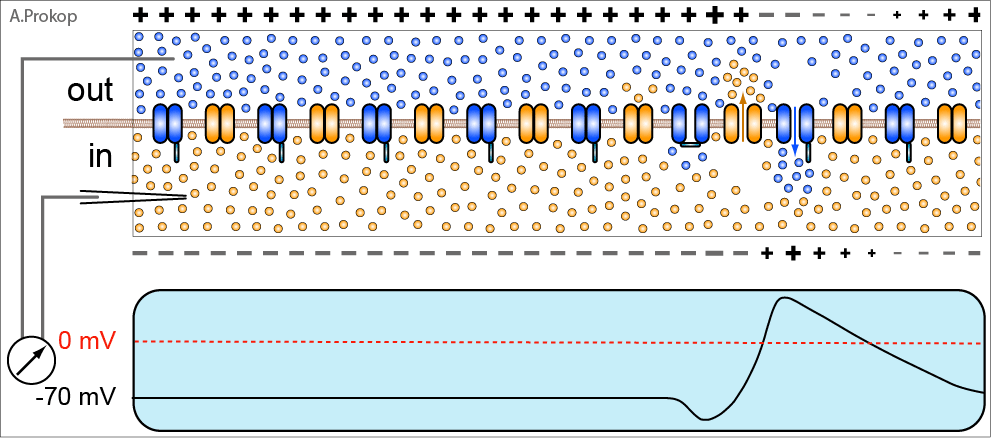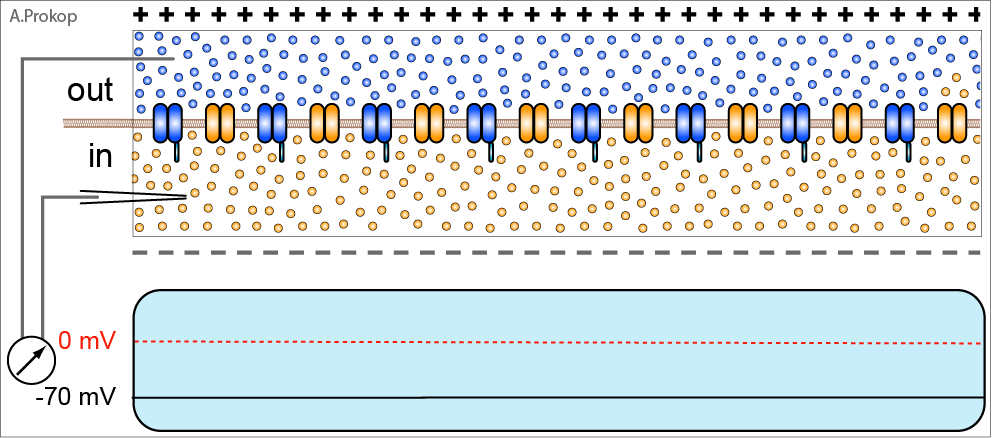
Рассмотрим синапс, образованный пресинаптической мембраной глутаматергического нейрона, т.е. выделяющего основной возбуждающий нейромедиатор глутамат, и постсинаптической мембраной нервной клетки, содержащей глутаматные рецепторы, которые обеспечивают проведение нервного сигнала.
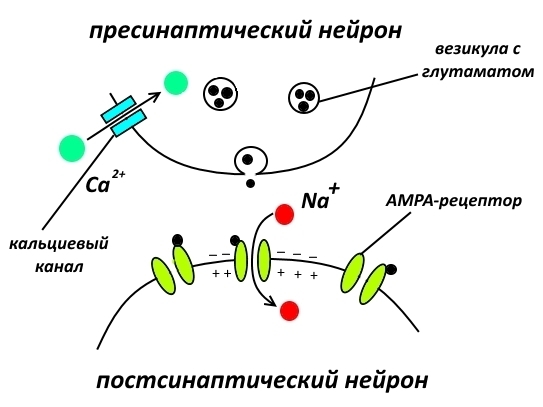
В стандартной ситуации нервный импульс проводится следующим образом (описание упрощено). Кальциевые каналы, расположенные в окончании пресинаптического нейрона, открываются, ионы кальция поступают внутрь и вызывают выделение нейромедиатора из везикул в синаптическую щель. Глутамат связывается с AMPA-рецепторами, ионный канал в них открывается, ионы натрия входят внутрь, вызывая деполяризацию. Возникающая деполяризация приводит к каскаду чередующихся закрытий/открытий ионных (калиевых и натриевых) каналов по аксону постсинаптического нейрона, направляя полученный сигнал к следующему синапсу. Усё.
Ситуация меняется, если постсинаптический нейрон получает значимый сигнал, т.е. когда происходит либо многократный выброс медиатора в синаптическую щель, либо выброс его очень большой дозы.
Вспомните, как вы учили стихи в школе – путём многократного проговаривания строчек. Так вы формировали значимый нервный сигнал и, сами того не подозревая, способствовали возникновению в определённых частях своего мозга долговременной потенциации – явления, которое, как полагают, и лежит в основе памяти и о котором далее пойдёт речь. Именно долговременная потенциация способствует укреплению связей между нейронами.
В электрофизиологических экспериментах с нейронами долговременную потенциацию вызывают тетанизацией – подачей через возбуждающий электрод короткого (1 с) высокочастотного (100 Гц) импульса.
Если полученный постсинаптическим нейроном сигнал был значимым, то деполяризация, т.е. обращение полярности обкладок конденсатора, образованных постсинаптической мембраной, оказывается сильнее, распространяется по постсинаптической мембране на большее расстояние и начинает действовать на другой тип глутаматных рецепторов – NMDA-рецепторы, расположенные рядом с AMPA-рецепторами.
NMDA-рецепторы тоже реагируют на глутамат, но одного глутамата им мало. Их ионный канал забит магниевой заглушкой. И снять её может лишь сильная деполяризация, возникающая при получении постсинаптическим нейроном значимого сигнала.
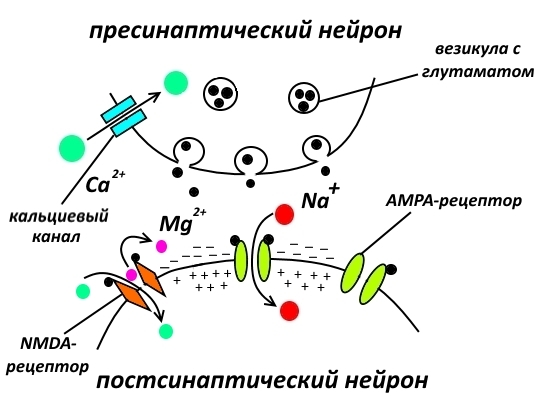
Снятие заглушки и одновременное связывание глутамата вызывает ток ионов через канал NMDA-рецептора. Но что это за ток? Наряду с ионами натрия в клетку начинают устремляться и ионы кальция, для которых ионный канал NMDA-рецептора также проницаем. А ионы кальция выполняют в нервной системе (да и вообще в организме) очень важную регуляторную функцию, вот почему его концентрация в органеллах клетки строго контролируется.
Ионы кальция действуют путём активации кальций-зависимых киназ – ферментов, фосфорилирующих различные белки (о них я уже писал немножко тута).
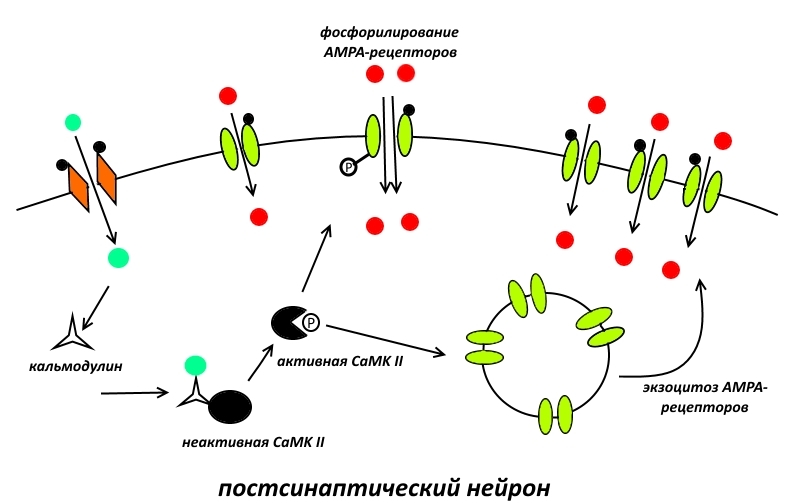
Первой на сцену выходит кальций/кальмодулин-зависимая протеинкиназа II (CaMK II). Она делает 2 важных вещи:
— фосфорилирует AMPA-рецепторы по остатку серина. Это приводит к увеличению их натриевой проводимости;
— вызывает экзоцитоз AMPA-из цитоплазмы на поверхность постсинаптической мембраны.
Таким образом, CaMKII усиливает сигнал, получаемый постсинаптическим нейроном за счёт увеличения числа и активности его приёмников – AMPA-рецепторов.
Поступление ионов кальция повышает в постсинаптическом нейроне активность адентилатциклазы – фермента, синтезирующего так называемый вторичный посредник – циклический аденозинмонофосфат, cAMP. сAMP активирует вторую киназу – протеинкиназу А (PKA), которая в постсинаптическом нейроне одно очень важное дело – проникает в ядро и фосфорилирует молекулу с названием из очередного набора ничего не говорящих слов – CREB (cAMP response element-binding protein) – транскрипционный фактор, активирующий (с помощью других кофакторов) транскрипцию и синтез некоторых белков. Про транскрипционные факторы я тоже немного писал.

«Что же это за белки?» — спросят те, кто смог дочитать до этого места. Отвечаю: современная наука ещё только-только приступила к определению их природы. Могу сказать о роли одного из них – BDNF (Brain-Derived Neurotrophic Factor), или нейротрофическом факторе роста.
Оказалось, что постсинаптический нейрон, задействованный в процессе долговременной потенциации, увеличивает площадь синапса, а также число синапсов, связывающих его с нейроном пресинаптическим. Естественно, всё это приводит к усилению (упрочнению и ускорению) его связи с пресинаптическим нейроном. За этот процесс отвечает как раз нейротрофический фактор роста BDNF.
Итак, резюмирую.
Явление долговременной потенциации, лежащее, как полагают, в процессе формирования долговременной памяти, состоит в усилении связи между взаимодействующими нейронами при прохождении через их синапс значимых сигналов. Такое усиление на ранних стадиях реализуется увеличением числа и проводимости AMPA-рецепторов по механизму, не зависящему от синтеза белков, а на поздних стадиях – увеличением числа и площади синапсов, связывающих нейроны. Поздние стадии долговременной потенциации требуют активации транскрипционных факторов и синтеза белка.
Я специально опустил некоторые важные детали, к которым хочу вернуться позднее, но многообразие белковых молекул, требующихся вам, чтобы разобраться в моей писанине и хоть что-то запомнить, должно быть заметно всем.
Внимание вопрос. Уважаемые скептики, неужели вам неочевидно, что изучение механизмов формирования памяти открывает весьма широкие возможности по созданию низкомолекулярных модуляторов (читай – ноотропных препаратов), которые позволят модифицировать её в нужном направлении?
Спасибо за внимание. Продолжение следует.
ссылка на оригинал статьи https://geektimes.ru/post/274811/
Добавить комментарий